Fly brain majorly composes three parts. Ventral neural cord (studied during embryo), medulla inside optic lobe and type ii neuroblast generating adult central complex (stuied during larva and puapl). Each of them has beautiful story to represent stem cell neural commitment mechanism.

Shows the temporal transcriptional factor (tTF) cascade in these three parts. Note that they apply overlapped though different tTF for their regulation, indicating only the basical principle (development needs temporal regulation) was shared even inside fly. Also, prior tTF activate next tTF, whereas is latter suppressed by next tTF is another fact that shared in various context.

There are 3 types of neuroblast exist in fly CNS.
Type I neuroblasts (the vast majority type inside neuroblasts) pass through self-renewing asymmetric divisions, producing a ganglion mother cell (GMC), which divides once more to generate a pair of neurons or glia (Note that the spatial localization of neuroepithelium, NM and GMC, follow a beautiful sequence based on differentiation degree).
Type II neuroblast (only 8 cells in larva brain lobe during development, responsible for generating adult central complex) reminisent oRG in mamaliam brain, which produce intermediate nneural progenitors (INPs). Each INP undergoes several asymmetric divisions, and each division generates a GMC that produces neurons or glia. Both neuroblasts and INPs progress through their own tTF series in parallel.
Type 0 (rare and only observe at a subset of neuroblasts) is the result of self-renewing asymmetric divisions that produce a neuroblast (NB) and a single neuron (become major in vertebrate). It occurs during the Dll temporal window (medulla tTF cascade), neuroblasts divide asymmetrically to self-renew and generate a neuron directly before switching to type I neurogenesis for the remainder of the temporal series.
Note all neuroblasts undergo tTF cascade whereas their progeny can be different, suggesting temporal regulation only roughly divide cell types (like generate different unicolumn neurons that ubiquitously distrbute all 800 colum in medulla) whereas additional regulation such as spatial pattern (generate spatial restricted multicolumn neurons) and Notch on/off (decide neurotransmitter type) further fine tuning and togetherly create the neuron reperoite.
And these three will be detailed discussed in the next passage.
1, Ventral neural cord (VNC)
https://www.sciencedirect.com/science/article/pii/S0092867401004652?via%3Dihub
Cell, 2001
The Drosophila VNC is analogous to the vertebrate spinal cord. During embryogenesis, neuroblasts with unique spatial identities, defined by their segmentally repeated bilateral organization, give rise to all neurons and glia of the VNC.
As they age, individual neuroblasts of the VNC sequentially express a series of transcription factors (known as temporal Transcription Factors or tTFs). The temporally restricted expression of these transcription factors defines unique identity windows during which different neurons are generated. Although the neuroblast itself changes tTF expression over time, neurons born within a given tTF window usually maintain expression of that tTF. Most VNC neuroblasts express the tTF series: Hunchback (Hb) → Krüppel (Kr) → POU domain proteins 1 and 2 (referred to as ‘Pdm’) → Castor (Cas) → Grainyhead (Grh) (Figure 1F) [6,7]. Neurons adopt distinct fates based on the tTF window in which they were born (it solely can not decide neural type, e.g., Hb+ can have multiple types which are distinguished by their projection, morpology and aborization pattern. But still, Hb+ positive neurons are firstly borned in progeny, compared to Kr or Pdm+ neurons, shows the tTF cascade decide the time sequence of neural commitment).
2, Medulla
Development, 2011
The optic lobes constitute two-thirds of the fly brain, and each of their neuropils—the lamina, medulla, lobula, and lobula plate —is divided into ~800 columns, corresponding to the same number of ommatidia (unit eyes) in the retina.
The medulla is the largest component of the Drosophila optic lobe, containing ~40,000 neurons (fly has 100,000 neurons) forming 10 layers (consider fly central brain is a intermingle netruophil, medulla is the most arranged structure in fly CNS). In Golgi studies, 60 types (another paper said 80 types) of medulla neurons with different morphological characteristics have been identified. These 80 types of neurons belong to two classes: uni-columnar neurons have a stoichiometry of one per column (each type has exactly 800 neurons, distributed one for each column. The cell body reside at the most distal part of medulla), while multi-columnar neurons contact multiple columns (fewer than 800, but has larger receptive field, bigger aborization, cover multiple column, and their cell body is proximal to medial medulla).

Here all the labeled neurons are uni-columnar neurons.
The medulla develops from a crescent-shaped neuroepithelium, the outer proliferation centre (OPC). During the third larval instar, the OPC neuroepithelium is converted into lamina on its lateral side and into medulla neuroblasts on its medial side. A wave of neurogenesis moves through the neuroepithelial cells, transforming them into neuroblasts; the youngest neuroblasts are closest to the neuroepithelium while the oldest are adjacent to the central brain.

Hth is expressed the eariliest and pushed inward by latter born Run+ and Drf+ neurons. Therefore, these concentric pattern conincidence with temproal transcriptional factor expressing pattern, since fly applies out-inside neuron production mode. (neuroblast resides at the outter most region, called OPC, so reverse plus reverse equal the same pattern of veterbrate. Early born neuron at innner and late born neuron at outside)
the developing medulla is subdivided into concentric zones that are characterized by the expression of four conserved transcription factors: Drifter (Drf), Runt (Run), Homothorax (Hth) and Brain-specific homeobox (Bsh). The expression patterns of these factors correlate with the order of neuron production. Once the concentric zones are specified, the distribution of medulla neurons changes rapidly, and the concentric zones are disrupted early in pupal development owing to the extensive migration of cell bodies.
Such tTF sequence has been further identified by Nature 2013 from Claude Desplan lab (he made all the works in this field). As the figure shown below.
https://www.nature.com/articles/nature12319
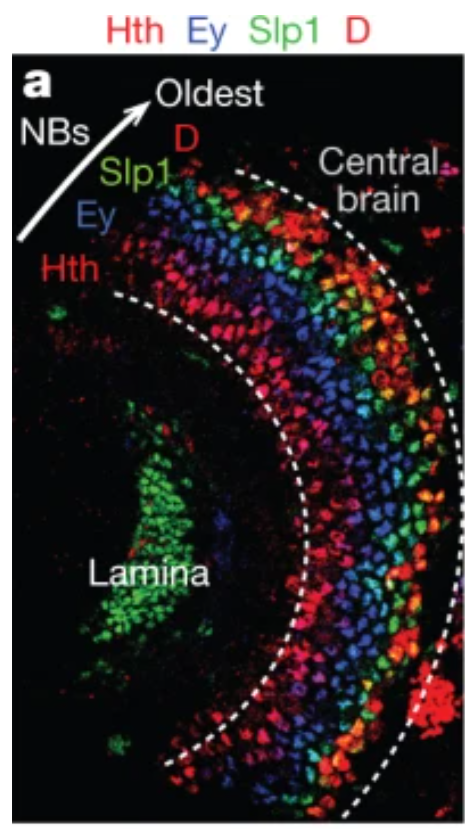

Left is the IHC of Hth-Ey-Slp-D tTF sequence. Right is the most clear diagram of medulla development I have ever seen. NB inside the OPC are all produced by medial NE, make the medial-lateral axis (lateral NBs are the oldest with biggest progeny at given time). tTF are coincident with dorsal-ventral axis, where oldest GMC (Hth+) stacked near the neuropil and yonger GMC at the distal region. This is the real reason that why concentric genes patterns are found.
3, Type II neuroblast INP
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3941985/#R20
Nature, 2013
Drosophila INPs sequentially generate distinct neural subtypes; that INPs sequentially express Dichaete>Grainyhead>Eyeless transcription factors; and that these transcription factors are required for the production of distinct neural subtypes. Moreover, parental type II neuroblasts also sequentially express transcription factors and generate different neuronal/glial progeny over time, providing a second temporal identity axis. We conclude that neuroblast and INP temporal patterning axes act combinatorially to generate increased neural diversity within adult central complex.


The green cluster is a clone. Fly cell do not move too much (in this case), means from left to right are the cells borned late to early (early cells been pushed outward). Thus it can provide insights that eyeless (homolog of PAX6 in vertebrate) is expressed latest.